Živčana stanica (neuron) proizvodi i provodi akcijski potencijal duž aksona, zatim prenosi signal preko sinapse otpuštajući neuroprijenosnike (neurotransmitere) što aktivira reakciju u drugom neuronu ili ciljnoj stanici (npr. mišićna stanica, većina egzokrinih i endokrinih stanica). Neurotransmiteri omogućuju neuronima da međusobno komuniciraju. Neurotransmiteri koji se oslobađaju vežu se na receptore na drugom neuronu. Neuroni koji oslobađaju neurotransmitere nazivaju se presinaptički neuroni. Neuroni koji primaju signale neurotransmitera nazivaju se postsinaptički neuroni. Signal može stimulirati ili inhibirati ciljnu stanicu, ovisno o neuroprijenosnicima i receptorima na koje djeluje. Drugi čimbenici, uključujući lijekove i poremećaje, utječu na komunikaciju između neurona modulacijom proizvodnje i djelovanja neurotransmitera, uključujući
-
Njihovo oslobađanje, ponovno preuzimanje i razgradnja
-
Broj i funkcija postsinaptičkih neurotransmiterskih receptora
Ponekad se signali između neurona javljaju u obrnutom smjeru (naziva se retrogradna neurotransmisija). U takvim slučajevima, dendriti (prijamne grane neurona) na postsinaptičkim neuronima oslobađaju neurotransmitere koji utječu na receptore na presinaptičkim neuronima. Retrogradni prijenos može spriječiti presinaptičke neurone u otpuštanju dodatnih neurotransmitera i pomoći u kontroli razine aktivnosti i komunikacije među neuronima.
Živčani impuls
U SŽS–u, međusobne sveze su složene. Impuls s jednog neurona na drugi može prijeći iz
Neuron istodobno prima mnogo impulsa (ekscitirajuće i inhibirajuće) od drugih neurona i integrira ih u različite obrasce izbijanja.
Provođenje
Širenje (provođenje) akcijskog potencijala duž aksona je električno, uzrokovano promjenama iona natrija i kalija na membrani aksona. Pojedini neuron proizvodi identični akcijski potencijal nakon svakog podražaja, provodeći ga konstantnom brzinom duž aksona. Brzina ovisi o promjeru aksona te stupnju mijelinizacije a kreće se od 1 do 4 m/s u malih nemijeliniziranih vlakana do 75 m/s u velikih mijeliniziranih vlakana. Brzina provođenja je veća u mijeliniziranih vlakana jer mijelinska ovojnica ima pravilno raspoređene pukotine (Ranvierovi čvorići) gdje je akson ogoljen. Električni impulsi skaču s jednog čvorića na sljedeći preskačući mijelinizirane dijelove aksona. Dakle, poremećaji koji mijenjaju mijelinski omotač (npr. multipla skleroza, Guillain-Barré sindrom) ometaju širenje impulsa, uzrokujući tako razne neurološke simptome.
Prijenos
Prijenos akcijskog potencijala je kemijska reakcija uzrokovana otpuštanjem specifičnih neuroprijenosnika iz živčanih završetaka aksona (terminala). Neuroprijenosnici difuzijom prelaze sinaptičku pukotinu te se kratkotrajno vežu za specifične receptore na susjednom neuronu ili ciljnoj stanici. Ovisno o receptoru odgovor može biti ekscitacijski ili inhibicijski. Obično se neuroni ne dodiruju; umjesto toga, oni komuniciraju putem prijenosa neurotransmitera preko sinapsi. Pod nekim uvjetima, neuroni koji su blizu jedan drugoga mogu komunicirati pomoću električnih impulsa preko praznine.
Tijelo živčane stanice proizvodi enzime koji sintetiziraju većinu neurotransmitera koji su pohranjeni u vezikulama na kraju neurona (pogledaj sliku Neurotransmisija). Količina neurotransmitera u jednoj vezikuli (najčešće više tisuća molekula) naziva se kvant. Akcijski potencijal po dolasku na terminal otvara aksonalne kalcijske kanale, utok kalcija u akson dovodi do spajanja membrane vezikula s membranom aksona što pak oslobađa neuroprijenosnike iz vezikula. To spajanje membrana stvara otvore kroz koje molekule bivaju egzocitozom izbačene u sinaptičku pukotinu.
Jedna vrsta sinapse, električna sinapsa, ne uključuje neurotransmitere; ionski kanali izravno povezuju citoplazme presinaptičkih i postsinaptičkih neurona. Ovakav je prijenos akcijskog potencijala najbrži.
Ekscitacijski i inhibitorni signali
Reakcija potaknuta otpuštanjem neurotransmitera može ili pobuditi ili aktivirati postsinaptički neuron ili inhibirati ili blokirati njegovu aktivnost. Postsinaptički neuroni primaju signale više neurotransmitera i električne signale od mnogih neurona. Neuron primatelj na kraju zbraja ulaze, a ako se primi više ekscitacijskih signala, neuron se aktivira i šalje signale drugim neuronima. Ako je zbroj signala inhibicijski, neuron se ne aktivira i ne utječe na aktivnost drugih neurona. Ovo zbrajanje odgovora naziva se zbrajanje. Neurotransmiteri tako olakšavaju brzu komunikaciju između neurona mijenjajući aktiviranje akcijskog potencijala.
Drugi oblici zbrajanja uključuju
Prostorno zbrajanje: Kada se primi više impulsa na različitim lokacijama neurona, a neuron ih tada zbraja
Vremensko zbrajanje: Kada se impulsi prime unutar kratkog vremenskog razdoblja i zatim se zbroje
Da bi neuron generirao signal i aktivirao se, mora dosegnuti potencijalni prag. Potencijal praga nastaje neto povećanjem dotoka natrija u stanicu tijekom izmjene iona natrija i kalija. Kada dovoljno natrija uđe u stanicu, dosegnut je prag; kada se dosegne prag, aktivira se akcijski potencijal; putuje duž membrane neurona. Mora se dosegnuti prag da bi se stvorio akcijski potencijal.
NeurotransmisijaAkcijski potencijal otvara aksonalne kalcijske kanale (nije prikazano). Ca++ aktivira otpuštanje neurotransmitera (NT) iz vezikula gdje su smješteni. Molekule NT ispunjavaju sinaptičku pukotinu. Neke se vežu za postsinaptičke receptore te izazivaju odgovor. Ostale bivaju ponovno uvučene i pohranjene u akson ili difundiraju u okolno tkivo. 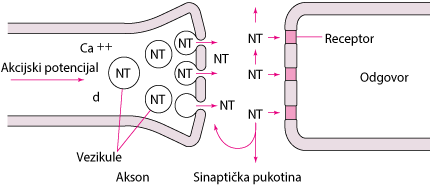 |
Količina neuroprijenosnika u živčanim završecima najčešće je neovisna o aktivnosti samog živca te se održava relativno konstantnom mijenjanjem unosa prekursora neuroprijenosnika ili promjenom aktivnosti enzima koji sudjeluju u sintezi neuroprijenosnika. Stimulacija presinaptičkih receptora može smanjiti sintezu presinaptičkih neuroprijenosnika dok ju blokada receptora može povećati.
Interakcija neuroprijenosnika i receptora mora se završiti brzo tako da se isti receptori mogu prestati kontinuirano aktivirati ili brzo ponovno aktivirati. S neurotransmiterima koji su stupili u interakciju s receptorima mogu se dogoditi sljedeće stvari:
Mogu se brzo prebaciti natrag u presinaptički terminal živca pomoću aktivnih procesa ovisnih o ATP (ponovna pohrana) za ponovni ciklus ili uništenje.
Mogu ih uništiti enzimi u blizini receptora.
Mogu difundirati u okolno područje i biti uklonjeni.
Neurotransmiteri koji su ponovno uneseni u aksonalni terminal opet ulaze u vezikule ili granule za naredni ciklus.
Poremećaji ovih procesa može rezultirati različitim bolestima. Na primjer, pretpostavlja se da gubitak pamćenja kod Alzheimerove bolesti uključuje nedostatak neurotransmitera acetilkolina u sinapsama, koji posreduje u polaganju novih sjećanja. Određeni lijekovi (npr. donepezil, galantamin, rivastigmin) blokiraju enzim acetilkolinesterazu (koji razgrađuje acetilkolin) i tako povećavaju količinu acetilkolina u sinapsi. Kao rezultat, pamćenje se može poboljšati.
Neke vrste pojedinačnih neurona mogu otpuštati dva ili više različitih neurotransmitera (koji se nazivaju kotransmisija)—na primjer, acetilkolin i glutamat. Više neurotransmitera može djelovati na jedan postsinaptički neuron ili utjecati na više postsinaptičkih neurona. Kotransmisija omogućuje zamršenu komunikaciju između neurona za kontrolu različitih događaja u CNS-u i perifernom živčanom sustavu (PNS).
Neurotransmiteri također mogu olakšati dugoročnije promjene koje uključuju dodatne putove kao što su promjene u aktivnosti gena i proteina.
Receptori:
Neurotransmiterski receptori su složeni proteini koji se protežu kroz staničnu membranu. Njihova vrsta određuje je li određeni neurotransmiter ekscitacijski ili inhibicijski. Receptori koji se kontinuirano stimuliraju neurotransmiterima ili lijekovima postaju neosjetljivi (regulirani na niže); oni koji nisu stimulirani svojim neurotransmiterima ili su kronično blokirani lijekovima postaju preosjetljivima (regulirani na više). Pozitivna i negativna povratna sprega ima jak utjecaj na razvoj tolerancije i fizičke ovisnosti. Ova spoznaja je izuzetno bitna kod transplantacije tkiva i organa gdje denervacija uskraćuje receptorima neurotransmitere, kao rezultat, transplantirani organ može postati preosjetljiv na živčanu stimulaciju. Simptomi ustezanja se mogu barem djelomično objasniti fenomenom odvikavanja uzrokovanog promjenom afiniteta i gustoće receptora.
Većina neuroprijenosnika primarno djeluje na postsinaptičke receptore, ali poneki se receptori nalaze na presinaptičkim neuronima te vrše kontrolu otpuštanja neuroprijenosnika.
Jedna porodica receptora, nazvan ionotropni receptori (npr, N-metil-d-glutamate, kinat-kviskvalat, nikotinski acetilkolin, glicin i gama-aminomaslačna kiselina [GABA] receptori), sastoje se od ionskih kanala koji se otvaraju vezanjem na neurotransmiter i izvršavaju vrlo brzi odgovor. U drugoj obitelji, nazvanoj metabotropnim receptorima (npr serotoninski, alfa- i beta-adrenergički i dopaminski receptori), neurotransmiteri stupaju u interakciju s G proteinima i aktiviraju drugu molekulu (drugi glasnik kao što je cAMP), koja katalizira lanac događaja putem fosforilacije proteina, mobilizacije kalcija, ili oboje. Stanične promjene koje posreduje drugi glasnik su sporije i dopuštaju finije podešavanje od brzog odgovora preko ionotropnih neurotransmitera. Daleko je više neurotransmitera koji aktiviraju specifične receptore nego druge glasnike.
Glavni neuroprijenosnici i receptori:
Postoji najmanje sto tvari koje mogu djelovati kao neurotransmiteri, od kojih je 18 vrlo značajno Nekolicina ih se javlja u malo drugačijim oblicima. Neurotransmiteri se mogu grupirati u različite klase, kao što su
Male molekule (npr. glutamat, gama-aminomaslačna kiselina, glicin, adenozin, acetilkolin, serotonin, histamin, noradrenalin)
Neuropeptidi (npr. endorfini)
Plinovite molekule (npr. dušikov oksid, ugljikov monoksid)
Endokanabinoidi
glutamat i aspartat:
Ove aminokiseline (glutamat i aspartat) su glavni ekscitacijski neuroprijenosnici u SŽS–u. Nalaze se u korteksu, malom mozgu i kralježničnoj moždini. Glutamat neurone potiče na povećanu sintezu dušičnog oksida (NO). Višak glutamata može biti toksičan povisujući intracelularni kalcij, slobodne radikale i aktivnost proteinaza. Ovi neurotransmiteri mogu doprinijeti toleranciji na opioide te posredovati hiperalgeziju.
Glutamatni receptori (stimulirani glutamatom i manje jakim aspartatom) su klasificirani kao NMDA (N–metil–D–aspartat) receptori i kao ne–NMDA receptori. Fenciklidin (PCP, poznat kao “anđeoski prah”) i memantin (koristi se za liječenje Alzheimerove bolesti) se vežu na NMDA receptore.
Gama-aminomaslačna kiselina
Gama-aminomaslačna kiselina GABA je glavni inhibitorni neuroprijenosnik u mozgu. Radi se o aminokiselini koja se stvara iz glutaminske kiseline dekarboksiliracijom uz pomoć glutamat dekarboksilaze. Nakon interakcije s receptorima, GABA se aktivnim transportom prenosi natrag u živčane završetke gdje se metabolizira. Glicin, koji je po funkciji sličan GABA, se javlja uglavnom u interneuronima (Renshawove stanice) kralježnične moždine te krugovima koji relaksiraju mišiće antagoniste.
GABA receptori su klasificirani kao GABA A (aktiviraju kloridne kanale) i GABA-B (pojačavaju stvaranje cAMP). GABAA receptori su mjesta djelovanja nekih neuroaktivnih lijekova uključujući benzodijazepine, barbiturate, pikrotoksin i muscimol. Alkohol se također veže na GABA-A receptore. GABAB receptore aktivira baklofen, koji se koristi u terapiji mišićnog spazma (npr. u multiploj sklerozi)
serotonin
Serotonin (5–hidroksitriptamin ili 5–HT) se proizvodi u nuclei raphes te neuronima ponsa i gornjeg dijela moždanog debla. Triptofan se hidroksilira uz pomoć triptofan hidroksilaze u 5–hidroksitriptofan, zatim se dekarboksilira u serotonin. Razina serotonina regulira se unosom triptofana i intraneuronalnom monoaminoksidazom (MAO), koja razgrađuje serotonin. U konačnici, serotonin se izlučuje u urinu kao 5-hidroksiindoacetat ili 5-HIAA.
Serotoninski (5-HT) receptori (s najmanje 15 podtipova) klasificirani su kao 5-HT1 (S 4 podtipa), 5-HT2, 5-HT3. Selektivni agonisti receptora serotonina (npr sumatriptan) mogu prekinuti migrene. Selektivni inhibitori ponovne pohrane serotonina (SSRI) također se mogu koristiti za liječenje nekoliko poremećaja mentalnog zdravlja (npr. depresija, anksioznost, opsesivno kompulzivni poremećaj, posttraumatski stresni poremećaj).
acetilkolin
Acetilkolin je glavni neurotransmiter bulbospinalnih motornih neurona, autonomnih preganglionskih vlakana, postganglionskih kolinergičkih (parasimpatičkih) vlakana, i mnogih neurona u središnjem živčanom sustavu (primjerice, u bazalnim ganglijima, motornom korteksu). Sintetizira se iz kolina i acetil koenzima A uz pomoć kolin acetiltransferaze, a njegovo djelovanje se brzo prekida lokalnom hidrolizom u kolin i acetat pomoću acetilkolinesteraze. Razine acetilkolina regulirane su kolin acetiltransferazom i ponovnom pohranom kolina. Razine ovog neurotransmitera su smanjene kod pacijenata s Alzheimerovom bolesti.
Kolinergički receptori su klasificirani kao nikotinski N1 (u srži nadbubrežne žlijezde i autonomnim ganglijima) i N2 (u skeletnim mišićima) ili muskarinski M1 do M5 (prisutan posvuda u živčanom sustavu). M1 se javlja u autonomnom živčanom sustavu, striatumu, korteksu i hipokampusu; M2 se javlja u autonomnom živčanom sustavu, srcu, crijevnim glatkim mišićima, te u strukturama stražnje lubanjske jame.
Dopamin stupa u interakciju s receptorima na nekim perifernim živčanim vlaknima i mnogim centralnim neuronima (npr. u substanciji nigri, srednjem mozgu, ventralnom tegmentalnom području i hipotalamusu). Aminokiselinu tirozin preuzimaju dopaminergički neuroni i pretvaraju je, pomoću tirozin hidroksilaze, u 3,4-dihidroksifenilalanin (dopa), koji se dekarboksilira u dopamin. Nakon otpuštanja i interakcije s receptorima, dopamin se aktivno vraća natrag u nervni završetak. Tirozin hidroksilaza i MAO (koji razgrađuje dopamin) reguliraju razinu dopamina u živčanim završetcima.
Dopaminski receptori su podijeljeni od D1 do D5. D3 i D4 receptori imaju važnu ulogu u kontroli misli (ograničavanje negativnih simptoma shizofrenije); dok aktivacija D2 receptora kontrolira ekstrapiramidni sustav. Međutim, afinitet receptora ne predviđa funkcionalni odgovor (intrinzičnu aktivnost). Ropinirol, koji ima visoki afinitet za D3 receptore ima intrinzičnu aktivnost putem aktivacije D2 receptora.
Norepinefrin je neuroprijenosnik većine postganglijskih simpatičkih vlakana i mnogih centralnih živčanih stanica (npr. u lokusu caeruleusu i hipotalamusu). Prekursor tirozin se pretvara u dopamin, koji se hidroksilira dopaminskom beta-hidroksilazom u noradrenalin. Nakon otpuštanja i interakcije s receptorima, dio noradrenalina se degradira katekolO-metiltransferazom (COMT), a ostatak se aktivno vraća u živčani završetak gdje ga razgrađuje MAO. Tirozin hidroksilaza, dopaminska beta-hidroksilaza, i MAO reguliraju intraneuronsku razinu noradrenalina.
Adrenergički receptori su klasificirani kao alfa-1 (postsinaptički u simpatičkom sustavu), alfa-2 (presinaptički u simpatičkom sustavu i postsinaptički u mozgu), beza-1 (u srcu), ili beta-2 (u drugim simpatički inerviranim strukturama).
Endorfini i enkefalini
Endorfini i enkefalini su opioidi.
Endorfini su veliki polipeptidi koji aktiviraju mnoge centralne neurone (npr. u hipotalamusu, amigdali. talamusu i lokusu caeruleusu). Tijelo stanica sadrži veliki polipeptid, pro-opiomelanokortin, koji je prethodnica alfa-, beta- i gama-endorfina. Pro-opiomelanokortin se transportira niz akson i cijepa na dijelove; jedan je beta-endorfin, koji se nalazi u neuronima koji se projiciraju u periakveduktalnu sivu tvar, limbičke strukture, te velike neurone u mozgu koji sadrže katekolamin. Nakon oslobađanja i interakcije s receptorima, beta-endorfin se hidrolizira peptidazama.
Enkefalin uključuje met-enkefalin i leu-enkefalin, koji su mali polipeptidi prisutni u mnogim centralnim neuronima (npr. u globusu pallidusu, talamusu, kaudatu i centralnoj sivoj tvari). Njihova prethodnica, proenkefalin, formira u tijelu stanice, a zatim se cijepa određenim peptidazama u aktivne peptide. Te tvari se također nalaze u kralježničnoj moždini, gdje moduliraju signale boli. Neurotransmiteri signala boli u stražnjim rogovima kralježnične moždine su glutamat i tvar P. Enkefalini smanjuju količinu neurotransmitera koja se otpušta i hiperpolariziraju (čine više negativnom) postsinaptičku membranu, čime se smanjuje stvaranje akcijskih potencijala i osjet boli na razini postcentralne brazde. Nakon otpuštanja i interakcije s peptidergičkim receptorima, enkefalini se hidroliziraju u manje, neaktivne peptide i aminokiseline. Brza inaktivacija egzogenih enkefalina sprječava da ove tvari budu klinički korisne. Umjesto toga, kao analgetici se koriste stabilnije molekule (npr, morfin).
Endorfin-enkefalin (opioidni) receptori klasificiraju se kao μ1 i μ2 (utječu na senzomotoričku integraciju i analgeziju), δ1 i δ2 (utječu na motornu integraciju, kognitivne funkcije i analgeziju), te κ1, κ2, i κ3 (utječu na regulaciju ravnoteže vode, analgeziju, i unos hrane). Sigma receptori, trenutno klasificirani kao neopioidni i uglavnom lokalizirani u hipokampusu, vežu PCP. Novi podaci ukazuju na prisutnost mnogo više podtipova receptora, s farmakološkim posljedicama. Komponente molekularnih preteča receptora mogu biti preuređene tijekom sinteze receptora u svrhu proizvodnju nekoliko varijanti receptora (npr. 27 varijanta cijepanja μ opioidnog receptora). Također, dva receptora se mogu spojiti (dimerizirati) kako bi stvorili novi receptor.
Ostali neurotransmitori
Dinorfini su skupina od 7 peptida sličnih aminokiselinskih sljedova. Oni su, kao i enkefalini, opioidi.
Tvar P, peptid, nalazi se u središnjim neuronima (u habenuli, supstanciji nigri, bazalnim ganglijima, moždini i hipotalamusu), a vrlo je koncentrirana u ganglijima dorzalnog korijena. Njeno izlučivanje se aktivira intenzivnim aferentnim bolnim podražajima. Modulira neuronalnu reakciju na bol i raspoloženje; a modulira i mučninu i povraćanje kroz aktivaciju NK1A receptora koji su smješteni u moždanom deblu.
Dušikov oksid (NO) je labilni plin koji posreduje u mnogim živčanim procesima. Generira se iz arginina pomoću NO sintaze. Neurotransmiteri koje povećavaju intracelularnu razinu kalcija (npr. tvar P, glutamat, acetilkolin) stimuliraju sintezu NO u neuronima koji eksprimiraju NO-sintetazu. NO bi mogao biti unutarstanični glasnik; mogao bi difundirati izvan stanice u drugi neuron i proizvesti fiziološke odgovore (npr dugoročno pojačavanje [jačanje određenih presinaptičkih i postsinaptičkih odgovora-oblik učenja]) ili pojačavati glutamatnu (NMDA-receptor-posredovanu) neurotoksičnost (npr. u Parkinsonovoj bolesti, moždanom udaru, ili Alzheimerovoj bolesti). NO utječe na druge neurotransmitere (npr. GABA i acetilkolin) mijenjajući dotok kalcija u stanice kako bi se povećalo otpuštanje drugih neurotransmitera.
Dodatni plinoviti neurotransmiteri uključuju ugljikov monoksid (CO) i vodikov sulfid (H2S). Ovi transmiteri se proizvode u stanicama cijelog tijela (uključujući mozak). Endogeni CO nastaje iz metabolizma hema i može sudjelovati u procesima koji uključuju stvaranje vrućice, upalu, preživljavanje stanica i kontrolu širenja krvnih žila. Nekoliko enzima uključeno je u proizvodnju H2S, za koji se vjeruje da je neophodan mozgu za formiranje i zadržavanje sjećanja.
Tvari s manje potvrđenim ulogama u neurotransmisiji uključuju histamin, vazopresin, vazoaktivni intestinalni peptid, karnozin, bradikinin, kolecistokinin, bombezin, somatostatin, kortikotropin-otpuštajući faktor, neurotenzin, i moguće adenozin.
Endokanabinoidi su endogeni neurotransmiteri na bazi lipida koji moduliraju funkciju mozga, endokrinog i imunološkog sustava.
Poremećaji povezani s defektima u neurotransmisiji
Poremećaji ili tvari koji mijenjaju proizvodnju, otpuštanje, primanje, razgradnju ili ponovnu pohranu neuroprijenosnika ili koji mijenjaju broj i afinitet receptora ogu uzrokovatineurološke ili psihijatrijske simptome i uzrokovati bolest (vidi tablicu Primjer poremećaja povezanih s defektima u neurotransmisiji). Lijekovi koji modificiraju neurotransmisiju mogu ublažiti ili ukloniti te poremećaje (npr. Parkinsonovu bolest, depresiju).
Primjeri poremećaja povezanih s defektima u neurotransmisijiPoremećaj | Patofiziologija | Liječenje |
|---|
* Eaton-Lambert sindrom je paraneoplastički sindrom posredovan protutijelima koji se tipično javlja kod malostaničnog karcinoma pluća. Može postojati prije nego se tumor manifestira. |
CRF = kortikotropin (ACTH) -otpuštajući faktor; GABA = γaminomaslačna kiselina; 5-HT = serotonin; IVIG = IV imunoglobulin; MAO = monoaminooksidaza; MAO-B = MAO tipa B; NMDA = N-metil-d-aspartat; NO = dušični oksid; NSAID = nesteroidni antireumatici; PIP2 = fosfatidilinozitol 4,5-bisfosfat; SSRI = selektivni inhibitori ponovne pohrane serotonina. |
Neravnoteža neuroprijenosnika |
|---|
Alzheimerova bolest | Izvanstanični depoziti beta-amiloida, unutarstanični neurofibrilarni čvorići i senilni plakovi, osobito u limbičkom sustavu (npr. hipokampus), u asocijativnom području korteksa, te u neuronima koji sintetiziraju i koriste acetilkolin (primjerice, u Meynertovoj bazalnoj jezgri i njezinim širokim projekcijama u korteks) | Inhibitori kolinesteraze (donepezil, rivastigmin, galantamin) odgađaju sinaptičku razgradnju acetilkolina i tako umjereno poboljšavaju kognitivnu funkciju i pamćenje. Memantin, antagonist NMDA– receptora, može usporiti napredovanje bolesti i povećati autonomiju bolesnika Budući tretmani mogu uključivati lijekove koji povećavaju razine H2S u mozgu i tako poboljšavaju kratkoročno pamćenje ili utječu na razine NO u mozgu i tako poboljšavaju dugoročno pamćenje. |
Anksioznost | Može odražavati smanjenu aktivnost GABA, možda zbog neravnoteže endogenih inhibitora, stimulatora GABA receptora ili oboje Može uključivati neravnotežu noradrenalinskog i 5-HT odgovora | Benzodiazepini povećavaju vjerojatnost otvaranja kloridnih kanala moduliranih GABA putem aktivacije GABA-A receptora. SSRI su lijekovi izbora za dugotrajno liječenje, jer se na benzodiazepine može razviti tolerancija |
Bolesti iz spektra autizma | Moguća hiperserotonemija, koja se javlja u 30-50% autističnih osoba, bez dokaza o središnjim 5-HT abnormalnostima | SSRI i risperidon mogu biti korisni |
Ozljeda mozga | Ozljede (npr, trauma, hipoksija, dugotrajne napadaje) stimuliraju prekomjerno oslobađanje ekscitatornih neurotransmitera (npr glutamata) i akumulaciju intracelularnih kalcijevih iona, koji doprinose smrti neurona | U eksperimentalnim modelima ishemije i ozljede, blokatori kalcijevih kanala, glicin i stariji antagonisti NMDA-receptora (npr. dekstrometorfan, ketamin) mogu smanjiti opseg gubitka neurona, ali ti lijekovi nisu učinkoviti kod ljudi. |
Depresija | Kompleksne abnormalnosti u kolinergičkom, katekolaminergičkom (noradrenergičkom, dopaminergičnom) i serotoninskom (5-HT) prijenosu Moguća uključenost drugih hormona i neuropeptida (npr. tvari P, dopamin, acetilkolin, GABA) | Antidepresivi snižavaju učinkovitost receptora indirektno ili direktno inhibicijom ponovnog preuzimanja 5-HT-a (kao SSRI-ovi) i norepinefrina ili dopamina te blokadom MAO-a. Blokada 5-HT2A/2C (Tip 5-HT receptora koji je u izobilju u prefrontalnom području) može povećati učinkovitost SSRI (npr. trazodon). |
Poremećaji napadaja | Napadaji koji se sastoje od iznenadnog sinkronog visokofrekventnog aktiviranja lokaliziranih skupina neurona u određenim područjima mozga, možda uzrokovanih povećanom aktivnošću glutamata ili smanjenom aktivnošću GABA | Fenitoin, lamotrigin, karbamazepin, valproat, topiramat, kao i neki drugi antikonvulzivi (npr. zonisamid, okskarbazepin) stabiliziraju natrijeve kanaliće ovisne o naponu. Etosuksimid i gabapentin smanjuju određene tokove kalcija. Fenitoin također smanjuje prekomjerno oslobađanje neurotransmitera. Lamotrigin može smanjiti razinu glutamata i aspartata. Fenobarbital i benzodiazepini pojačavaju aktivaciju GABA preko utjecaja na kompleks GABA-A receptor-kloridni kanal. Tiagabin blokira glijalni unos GABA. Valproat povećava razinu GABA. Topiramat povećava djelovanje GABA. |
Huntingtonova bolest (koreja) | Značajna oštećenja neurona u korteksu i striatumu zbog poliglutaminske ekspanzije (kodiran CAG ponavljanjima) kao rezultat nenormalnog gena na kromosomu 4. (abnormalni gen prekomjerno proizvodi protein huntingtin, koji se moguće kombinira s molekulama koji dovode do prekomjerne stimulacije stanica ekscitatornim aminokiselinskim neurotransmiterima kao što je glutamat) | Ne postoji specifično liječenje, ali lijekovi koji blokiraju NMDA receptore mogu blokirati toksične učinke viška glutamata. GABA–mimetički lijekovi nisu učinkoviti |
Manija | Povećana aktivnost noreadrenalina i dopamina reducira 5-HT razinu i abnormalnu glutamatnu neurotransmisiju. | Litij je tradicionalni prvi izbor. Smanjuje oslobađanje noradrenalina i povećava sintezu 5-HT. Valproat i lamotrigin su korisni, možda kroz normalizaciju transmisije glutamatom. Topiramat blokira natrijeve kanale ovisne o naponu, povećava GABA aktivnost na neke podvrste GABAA receptora, antagonizira AMPA / kainat podtip receptora glutamata te inhibira enzim karboanhidrazu, osobito izoenzime II i IV. Za gabapentin se smatra se da se veže na alfa-2/delta podjedinicu (1 i 2) kalcijevih kanala ovisnih o naponu u SŽS. Karbamazepin i okskarbazepin stabiliziraju natrijeve kanale ovisne o naponu. |
Neuroleptički maligni sindrom | Blokada od dopamin (D2) receptore lijekovima (npr. antipsihotici, metilfenidat) ili naglim povlačenjem dopaminergičkog agonista, što rezultira ukočenošću mišića, vrućicom, promjenom mentalnog statusa i autonomnom nestabilnošću | Liječenje D2 agonistom (npr. bromokriptin) poništava poremećaj neurotransmisije. Drugi lijekovi koriste se prema potrebi (na primjer, dantrolen, izravan mišićni blokator, se koristi za blokiranje grčeva mišića). |
Bol | Ozljeda tkiva, koja uzrokuje otpuštanje supstance P i glutamata u stražnjem rogu leđne moždine i oslobađanje drugih makromolekula koje posreduju signale boli, kao što je CGRP (koji može dilatirati kranijalne krvne žile i tako uzrokovati migrenu) i neurokinin A i bradikinin, koji su lokalizirani u prvom redu u lamini II i IV leđne moždine Daljnje modulacija tih signala putem endorfina (u leđnoj moždini) i 5-HT i noradrenalina (u silaznim putevima koji nastaju u mozgu) | NSAIL inhibiraju sintezu prostaglandina selektivno (s inhibitorima COX-2 — npr. celekoksib, parekoksib) ili neselektivno (s inhibitorima COX-1 i -2 — npr. ibuprofen, naproksen) i smanjuju stvaranje impulsa boli. Opioidni analgetici (npr. morfij) aktiviraju endorfin–enkefalinske (mi, delta i kapa) receptore, time smanjujući prijenos bolnih impulsa Novi tretmani koji mogu blokirati CGRP receptore mogu liječiti bol u živčanom sustavu. Ovi tretmani mijenjaju signalizaciju CGRP-a i tako ublažavaju bol tijekom migrena; mogu se uzeti u obzir za liječenje boli lica, dijabetičkih neuropatija i boli nakon liječenja raka. |
Parkinsonizam | Inhibicija dopaminergičkog sustava zbog blokade dopaminergičkih receptora antipsihoticima | Antikolinergički lijekovi smanjuju kolinergičku aktivnost i uspostavljaju ravnotežu između kolinergičkog i dopaminergičkog sustava. |
Parkinsonova bolest | Gubitak dopaminergičkih neurona pars compacta u substantia nigra i drugim područjima, sa smanjenim razinama dopamin i metenkefalin, mijenjajući dopamin/acetilkolinske ravnoteže i rezultira pretjeranom strijatalnom acetilkolinskom aktivnošću | Levodopa dospijeva u sinaptičku pukotinu, preuzima je akson kroz presinaptičke crne neurone i dekarboksilira se u dopamin, koji se izlučuje u pukotinu za aktiviranje dendritika dopamin receptore. Amantadin povećava presinaptičko otpuštanje dopamina; dopaminski agonisti stimuliraju dopaminske receptore, iako se bromokriptin, pramipeksol i ropinirol vežu samo za D2, D3 i D4 podtipove dopaminskih receptora. Antikolinergici reduciraju aktivnost kolinergičkog sustava, uspostavljajući tako balans dopamina i acetilkolina. Inhibitori MAO–B sprječavaju ponovni unos dopamina, čim dovode do porasta njegove razine u sinaptičkoj pukotini. Selegilin, inhibitor MAO–B,sprječava razgradnju dopamina, čime produžuje odgovor na levodopu i posljedično omogućuje smanjenje doze karbidope/levodope Katekol O-metiltransferaza(COMT) inhibitori, također, inhibiraju raspad dopamina. |
Shizofrenija | Povećano presinaptičko oslobađanje, sinteza dopamin, osjetljivost ili gustoća postsinaptičkog dopamin receptore ili kombinaciju | Antipsihotici blokiraju dopamin receptore i smanjiti dopaminergičku prekomjernu aktivnost na normalu. Haloperidol ponajviše blokira D2 i D3 receptore (visoki afinitet), te D4 receptore (niski afinitet) u mezokortikalnom području. Klozapin ima visoki afinitet za D4 i 5–HT2 receptore, što upućuje na uključenost serotoninskog sustava u patogenezu shizofrenije i njen odgovor na liječenje. Klozapin nosi značajan rizik za leukopeniju. Olanzapin i risperidon, slično haloperidolu, također imaju visok afinitet za 5–HT2 i D2 receptore |
Tardivna diskinezija | Preosjetljivo dopamin receptora zbog kronične blokade antipsihoticima | Smanjenje doza antipsihotika može smanjiti preosjetljivost dopamin receptori; međutim, u nekim slučajevima, promjene mogu biti nepovratne. |
Normalni neurotransmiteri koje imunološki sustav blokira ili uništava |
|---|
Miastenija gravis | Odražava inaktivaciju acetilkolinskih receptora i postsinaptičke histokemijske promjene na neuromuskularnom spoju zbog autoimunih reakcija | Antikolinesterazni lijekovi inhibiraju acetilkolinesterazu, povećavaju razinu acetilkolina na spoju i stimuliraju preostale receptore, povećavajući aktivnost mišića. |
Smanjen ponovni unos neuroprijenosnika u živčane stanice |
|---|
Amiotrofična lateralna skleroza | Uništavanje gornjih i donjih motoričkih neurona, vjerojatno djelomično uzrokovano glutamatnom neurotoksičnošću | Riluzol, koji inhibira prijenos glutamata, umjereno produljuje preživljenje. |
Neuroprijenosnici normalni, ali oštećeni ionski kanali |
|---|
Epizodična ataksija | Neispravni naponski kontrolirani kalijevi kanali, uzrokujući distalno mreškanje i nekoordinaciju (miokimija) | Liječenje acetazolamidom je učinkovito u nekim vrstama epizodičkih ataksija. |
Hiperkalijemijska periodična paraliza | Smanjena inaktivacija natrijevih kanala | Teški napadi mogu se prekinuti kalcijevim glukonatom, glukozom i inzulinom. |
Hipokalemijska periodična paraliza | Neispravni naponski kalcijevi kanali | Akutni napadi mogu se prekinuti kalijevim solima. Acetazolamid je učinkovit za prevenciju |
Lambert-Eaton syndrome* | Antitijela koja smanjuju presinaptičko otpuštanje acetilkolina | Kortikosteroidi, 3,4-diaminopiridin (DAP), gvanidin, IVIG i plazmafereza mogu biti korisni. |
Urođena paramiotonija | Neispravni naponski kontrolirani natrijevi kanali, uzrokujući miotoniju izazvanu hladnoćom i epizodnu slabost | Meksiletin (blokator natrijevih kanala) i acetazolamid (inhibitor karboanhidraze) mogu biti korisni. |
Rasmussenov encefalitis | Postviralna proizvodnja antitijela na glutamatne receptora, što utječe na glutamat-ovisne kanale Najizrazitiji oblik epilepsije partialis continua | Kortikosteroidi i antivirusni lijekovi obično su neučinkoviti. Funkcionalnom hemisferektomijom (npr. rezanje corpusa callosuma) možemo kontrolirati napadaje ako ne dođe do spontane remisije bolesti |
Hiperekpleksija | Mutacija u genu za alfa-1 podjedinicu glicin-ovisnog kanala Karakterizira je ukočenost, noćni mioklonus i pretjeran "startle" refleks, s hiperrefleksijom i padovima | Klonazepam ili neki drugi antikonvulzivi (npr. fenitoin, fenobarbital, diazepam, valproat) mogu dovesti do poboljšanja. |
Trovanje |
|---|
Botulizam | Inhibicija otpuštanja acetilkolina iz motornih neurona toksinom iz Clostridium botulinum | Specifična terapija lijekovima ne postoji. Mala količina toksina se koristi za liječenje određenih distonija, spastičnosti, neuropatske boli i migrene, a kozmetički za smanjenje bora na koži. |
Otrovanje gljivama | Muhara: Sadrži ibotensku kiselinu (koja ima učinke slične onima glutamata) i metabolit sličan muscimolu (koji ima učinke slične onima GABA) Inocybe i Clitocybe spp: Stimulacija muskarinskih receptora muskarinom i srodnim spojevima | Liječenje je potporno, jer nema lijekova za preokretanje učinka na neurotransmisiju. Atropin pomaže zaustaviti muskarinske manifestacije. |
Organofosfati | Ireverzibilna inhibicija acetilkolinesteraze i izrazito povećanje razine acetilkolina u sinaptičkoj pukotini | Pralidoksim uklanja toksine s acetilkolinesteraze i pomaže zaustaviti nikotinske kao i muskarinske manifestacije. Atropin pomaže brzo zaustaviti muskarinske učinke. |
Otrov zmije Bungarus multicinctus | Blokira receptore acetilkolina na neuromuskularnom spoju alfa-Bungarus toksin | Protuotrov je dostupan i čini se učinkovitim. |